I always like the geo coins but maybe im slow . Are these just for collecting do they have a pourpose other then just being cool.
Geo coins with tracking numbers are meant to be placed in Geo caches for other geocachers to find. They then log there finds on Geocaching.com Here is the list of coins I have discovered. http://www.geocaching.com/profile/
There are thousands of different coins you can buy, some just collect them others sent them out for others to find and move on to other caches.
Here is one of mine that has traveled over 6300 miles and is now in Germany.
Not to hijack the thread but these were from the Tyler build. Does anyone need one? I would like to get one of these to someone who needs one.....not someone who already has one:
I found a website who makes metal ones but i'd rather buy from a fellow pennocks fiero owner geocacher.Pm me if you do and how much for about 20 as thats how many caches i own and like to drop a coin in.I understand if the order has to be 50 or more.jesse
Can anyone get a hold of race? He is the one that designed the coin. I just thought i would see if we could spark enough interest for race to make another coin
I found a website who makes metal ones but i'd rather buy from a fellow pennocks fiero owner geocacher.Pm me if you do and how much for about 20 as thats how many caches i own and like to drop a coin in.I understand if the order has to be 50 or more.jesse
The coins are really to nice to use as geocoins as I have lost all of the ones I sent out to bad cachers or mugglers.
Not to hijack the thread but these were from the Tyler build. Does anyone need one? I would like to get one of these to someone who needs one.....not someone who already has one:
I haven't got one of those. I didn't know there were a bunch made.
Originally posted by reborngt: Why dont we do one with a tribute to pontiac on one side and fiero on the other.
I dont think redesigning one of the coin's sides would be cost affective especially for copyright ...... I think we should do one for the Indy White and Red....
What was the last round? Was it the bare metal silver coin? That was the first, and only coin that I was able to get, I wish I knew about these before that because I would sure love to own one of each that has been made! If anyone has a spare or two they would like to get rid of send me a pm!
RACE, if you do end up making another batch, I would order three! Chris
Danyel is right about making new dies. I would have to send a prototype coin to EMI ans it would be cost prohibitive and possibly wouldn't even be approved without having to pay them. That would put the price out of reach for the average coin buyer. It has been my goal from the start to provide coins at the lowest price possible so that anyone who wants them can afford to own them on any budget.
I will consider the Indy coin. Possibly on a black nickel base coin to preserve the color scheme of the car. I am open to other ideas too.
Originally posted by RACE: I will consider the Indy coin. Possibly on a black nickel base coin to preserve the color scheme of the car. I am open to other ideas too.
I wanna second that !!! hope i'm still on your email list for these !!!!
I don't think that an Indy coin would be all that flashy but I guess it would be historically significant. I would like to match the red though. I'll see what people think about it.
I don't think that an Indy coin would be all that flashy but I guess it would be historically significant. I would like to match the red though. I'll see what people think about it.
Hey guys, just an FYI. I cannot locate the other coins that I have. I have received a lot of emails about the Tyler coin but cannot find the remaining 3 that I have so I cannot hand them out
The Indy is prominently white .... so I say it should be white in the back too... IMHO Danyel
I'm just a little concerned that a black white coin might not look all that good, even with a red ring on the front. If I could find my artwork file I could show you. I should be able to get it when I get to my laptop in Memphis later this week.
Not to hijack the thread but these were from the Tyler build. Does anyone need one? I would like to get one of these to someone who needs one.....not someone who already has one:
Ill start a thread to produce the Indy Geocoin and post a link here in a little bit once it is set up. Please make your official quantity and engraving requests on the new page so that I can keep track of everyone's requests more easily.
Here is the link for ordering your Indy Fiero Geocoins.
Edited to fix the link. Thanks
[This message has been edited by RACE (edited 04-20-2012).]
------------------ When I read about the evils of drinking. I gave up reading. 2004 White Sebring Convertible 1986 Black Fiero GT Kathryn's Welded Metal Artist
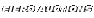
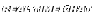






















 I could be talked into another round.
I could be talked into another round. 










 Thanks
Thanks




